

Environmental responsiveness and phenotypic plasticity are omnipresent in nature. All organisms are exposed to an environment and most of these environments are changing constantly, often in an unpredictable manner. Not surprisingly therefore, plasticity is found in all domains of life and at all levels of biological organization.
Developmental (phenotypic) plasticity describes the property of a genotype to respond to environmental variation by producing distinct phenotypes. The concept of plasticity dates back to the beginning of the 20th century and has continuously been developed to arrive at its current state, where many practitioners consider plasticity to represent a major facilitator of evolutionary diversification. In our lab, we investigate developmental plasticity at an integrative level.
WHAT IS OUR MODEL SYSTEM?
We are using the mouth-form dimorphism in Pristionchus pacificus and its relatives to study the questions mentioned above.
P. pacificus and related nematodes lives on scarab beetles, i.e. cockchafers and stag beetles. Pristionchus waits for the beetle to die before exiting the arrested dauer stage, a so-called necromenic association. On the beetle carcass, there is enormous competition for food and survival between many animals and microbes all living in this niche. It is long known that Pristionchus and relatives form teeth-like denticles in their mouth, which allow predatory feeding (Fig. 1). In the case of P. pacificus, animals decide during larval development in an irreversible manner to adopt a eurystomatous (Eu) or a stenostomatous (St) mouth-form. Eu animals form two teeth with a wide buccal cavity, representing predators. In contrast, St animals have a single tooth with a narrow buccal cavity and are strict microbial feeders. This dimorphism represents an example of phenotypic plasticity (Bento et al., 2010). Most importantly, mouth-form plasticity is regulated by conditional factors such as crowding, but also contains stochastic elements of regulation: a nearly constant ratio of 70-90% Eu : 30-10% St animals is formed under fixed environmental conditions. It is this aspect of stochastic regulation that allows manipulation of plasticity by genetic and molecular tools. Indeed, unbiased genetic screens resulted in the isolation of mutants that would only form one of the two mouth-forms and some of these mutants were characterized to represent “developmental switch genes”. For example, eud-1 mutants are monomorphic, being all-St (Ragsdale et al., 2013). Since these original findings, the gene regulatory network determining the P. pacificus mouth form has been studied in great detail (for recent reviews see Wighard and Sommer; 2025; Sommer, 2025).
Work on mouth-form plasticity builds on unique advantages of the Pristionchus system as model for plasticity research:
Given these unique assets, the major aim of our research is to obtain
A HOLISTIC UNDERSTANDING OF DEVELOPMENTAL PLASTICITY AS FACILITATOR OF PHENOTYPIC EVOLUTION
To achieve this goal, we perform three long-standing research directions in the Department:
Long-term environmental induction and experimental evolution
The elucidation of the genetic, epigenetic and other molecular processes underlying mouth-form plasticity provides, for the first time, a detailed mechanistic understanding of any plastic trait in animals. However, such studies themselves do not reveal if developmental plasticity is important for evolution.
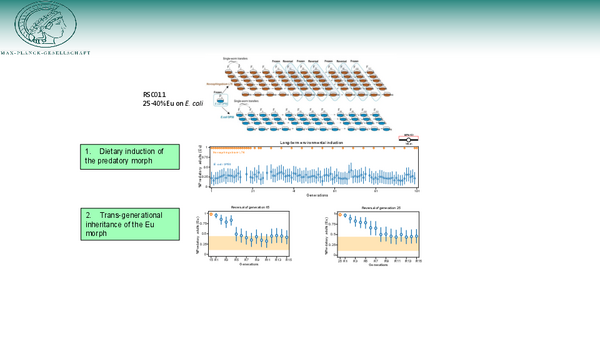
To obtain comprehensive evidence for the significance of developmental plasticity for evolution, we have started during the Corona pandemic, unprecedented long-term environmental induction experiments through experimental evolution (Fig. 2). Specifically, we investigated the interplay between feeding structure plasticity and the environment on an evolutionary time scale, by subjecting a total of 110 isogenic nematode lines to different bacterial diets (E. coli vs. Novosphingobium) for more than 100 generations (Fig. 2). This long-term evolution experiment illustrates i) systemic dietary induction of the predatory mouth form (Fig. 2), and ii) transgenerational epigenetic inheritance (TEI) of the predatory phenotype after food reversal (Fig. 2). In our case, TEI requires a dietary induction of at least five generations, which is distinct from previously described examples of TEI in C. elegans (see Area 2 below). These significant findings are at the center of our current work and the work that the Department will focus on in the coming years. They are described below:
AREA 1: Dietary induction of the predatory mouth-form and its regulation by vitamin B12
We used bacteria isolated from Pristionchus-associated environments to study dietary influences on mouth-form expression. We have originally found that a Novosphingobium diet causes immediate, systemic and complete induction of the predatory mouth form (Fig. 2). By now, we have identified vitamin B12 as the causal agent for the induction of the predatory morph and also its subsequent transgenerational transmission. This is the first demonstration of a vitamin in transgenerational inheritance. Our current work tries to explore the many relevant links from the 1-carbon cycle (methionine, vitamin B9/folate and SAM) to the role of vitamin B12 in neuropathologies.
AREA 2: Transgenerational inheritance of the predatory mouth form and microRNA signaling
To study this phenomenologically novel type of TEI (Fig. 2), we coupled experimental evolution with unbiased forward genetic analysis. We identified a role of the ubiquitin ligase EBAX-1/ZSWIM8 and, for the first time, of a microRNA for the transmission of transgenerational inheritance. Ppa-EBAX-1 destabilizes the microRNA family miR-2235, which underwent a massive expansion in P. pacificus relative to its C. elegans homolog miR-35. Through the generation of two deletion mutations of a cluster of 44 copies of miR-2235 that also harbors 42 interspersed copies of another microRNA family, we could provide the first evidence for miRNA signaling in transgenerational inheritance. These results are currently explored in many different directions.
In particular, saturation mutagenesis has resulted in the isolation of many novel candidate genes involved in TEI. These findings suggest an information flow from the intestine to the nervous system and from there to the germ line. Specifically, we have isolated a total of 165 independent mutations with a ‘transgenerational inheritance defective (tid) phenotype. Several scientists and technicians work in parallel on the most promising mutant candidates. In addition, we investigate the natural variation and evolution of the machinery associated with the transgenerational inheritance of the predatory morph. Importantly, there is no overlap in our candidates with the only other unbiased screen that was performed in C. elegans more than a decade ago and has largely found a role for the RNAi pathway being involved in transgenerational inheritance. We currently hypothesize that the much stronger role of miRNA signaling in the transgenerational inheritance of mouth-form plasticity in P. pacificus is responsible for this different spectrum of mutants that will allow us to provide fundamentally novel insight into transgenerational memory transition.
MACRO-EVOLUTIONARY POTENTIAL OF PLASTICITY AND FACILITATION OF DIVERSITY
Is plasticity important for evolutionary diversification and novelty? Answering this question requires comparative studies that when performed in a phylogenetic context can provide insight into the significance of plasticity for evolutionary processes. Two previous studies have moved this analysis to the macro-evolutionary level, suggesting that phenotypic plasticity indeed facilitates rapid diversification. First, we studied the evolution of feeding structures in more than 90 nematode species using geometric morphometrics (Susoy et al., 2015). This study found that feeding dimorphism was indeed associated with a strong increase in complexity of mouth-form structures. At the same time, the subsequent assimilation of a single mouth-form phenotype coincided with a decrease in morphological complexity, but an increase in evolutionary rates.
A second case of mouth-form plasticity increasing morphological diversification came from a striking example of fig-associated Pristionchus nematodes (Susoy et al., 2016). These nematodes form five distinct mouth-forms that occur in succession in developing fig synconia, thereby increasing the polyphenism from two to five distinct morphs. Additionally, the morphological diversity of these five morphs exceeds that of several higher taxa, although all five morphs are formed by the same species. These findings strongly support the facilitator hypothesis and they also indicate that ecological diversity can be maintained in the absence of genetic variation as all this diversity is seen within a single species and without associated speciation and radiation events.
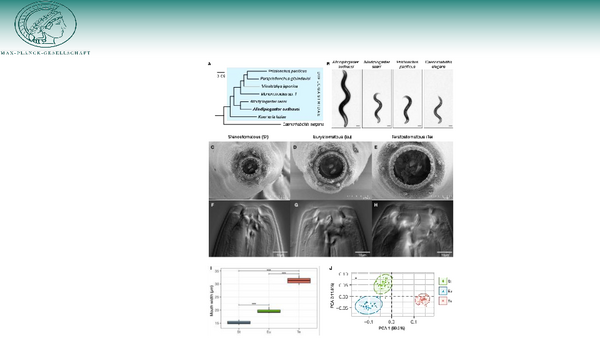
More recently, we started a functional analysis of mouth-form evolution using the distantly related diplogastrid Allodiplogaster sudhausi as model (Wighard et al., 2022) (Figure 3). This species form three distinct mouth forms, all of which are predators of other nematodes. Genetic studies showed that the gene regulatory network controlling mouth form plasticity is evolutionary conserved and eud-1-like sulfatases control feeding plasticity also in A. sudhausi (Wighard et al., 2024). A more detailed analysis of additional genes indicated conservation and change of the network throughout evolution (Wighard et al., 2025). These studies indicate the significant contribution of developmental plasticity for macroevolutionary divergence of important life history traits and highlight the short comings of mainstream evolutionary theory when neglecting the importance of developmental (phenotypic) plasticity.
Quiobe, S. P., Sommer, R. J. (2025). Adaptive consequences of transgenerational inheritance of a predatory mouth-form trait in nematodes. bioRxiv 2025.10.05.680509; doi: doi.org/10.1101/2025.10.05.680509
Dardiry, M., Piskobulu, V., Kalirad, A. & Sommer, R. J. (2023). Experimental and theoretical support for costs of plasticity and phenotype in a nematode cannibalistic trait. Evolution Letters, 7, 48-57.
Werner, M. S., Loschko, T., Franz-Wachtel, M., Theska, T., Macek, B. & R. J. Sommer (2023): Histone 4 lysine 5/12 acetylation enables developmental plasticity of Pristionchus mouth form. Nature Communications, 14: 2095
Lo, W.-S., Roca, M., Dardiry, M., Mackie, M., Eberhardt, G., Witte, H., Hong, R., Sommer, R. J. & J. W. Lightfoot (2022): Evolution and diversity of TGF-β signalling pathways are linked with novel developmental and behavioural traits. Molecular Biology and Evolution, 39: msac252.
Lightfoot, J. W., Dardiry, M., Kalirad, A., Giaimo, S., Eberhardt, G., Witte, H., Wilecki, M., Roedelsperger, C., Traulsen, A., & R. J. Sommer, R.J. (2021): Sex or cannibalism: polyphenism and kin recognition control social action startegies in nematodes. Science Advances, 7, eabg8042.
Theska, T., Sieriebriennikov, B., Wighard, S. S., Werner, M. S. & Sommer, R. J. (2020): Geometric morphometrics of microscopic animals as exemplified in model nematodes. Nature Protocols, 15, 2611-2644.
Sieriebriennikov, B., Sun S., Lightfoot, J.W., Witte, H., Moreno, E., Rödelsperger, C., Sommer, R.J. (2020): Conserved nuclear hormone receptors controlling a novel plastic trait target fast-evolving genes expressed in a single cell. PLOS Genetics. doi.org/10.1371/journal.pgen.1008687
Sommer, R.J. (2020): Phenotypic Plasticity: From Theory and Genetics to Current and Future Challenges. Genetics, Vol. 215, 1-13
Sieriebriennikov, B. & Sommer, R. J. (2018): Developmental plasticity and robustness of a nematode mouth-form polyphenism. Frontiers in Genetics. doi:10.3389/fgene.2018.00382
Sieriebriennikov, B., Prabh, N., Dardiry, M., Witte, H., Rödelsperger, C., Röseler, W., Kieninger, M. R. & Sommer, R. J. (2018): A developmental switch generating phenotypic plasticity is part of a conserved multi-gene locus. Cell Reports, 23, 2835-2843.
Serobyan, V., Xiao, H., Rödelsperger, C., Namdeo, S., Röseler, W., Witte, H. & Sommer, R. J. (2016): Chromatin remodeling and antisense-mediate up-regulation of the developmental switch gene eud-1 control predatory feeding plasticity. Nature Commun., 7: 12337.
Susoy, V., Herrmann, M., Kanzaki, N., Kruger, M., Nguyen, C.N., Rödelsperger, C., Röseler, W., Weiler, C., Giblin-Davis, R. M., Ragsdale, E. J. & Sommer, R. J. (2016): Large-scale diversification without genetic isolation in nematode symbionts of figs. Science Advance, 2: e1501031.
Susoy, V., Ragsdale, E. J., Kanzaki, N. & Sommer, R. J. (2015): Rapid diversification associated with a macroevolutionary pulse of developmental plasticity. eLIFE, 4: e05463.
Featured in: Nijhout, H.F. (2015): To plasticity and back again. eLIFE, 4: e06995.
Ragsdale, E. J., Mueller, M. R., Roedelsperger, C. & Sommer, R. J. (2013): A developmental switch coupled to the evolution of plasticity acts through a sulfatase. Cell, 155, 922-933.
Bento, G., Ogawa, A. & Sommer, R. J. (2010): Co-option of the hormone-signalling module dafachronic acid–DAF-12 in nematode evolution. Nature, 466, 494-497.
